Главная Поиск
Различные способы поиска
Поиск по базе данных:  Научные статьи Видеоматериалы
Научные статьи Видеоматериалы
 Поиск Яндексом по сайту
Поиск Яндексом по сайту
Репозиторий OAI—PMH
Репозиторий Российская Офтальмология Онлайн по протоколу OAI-PMH
Конференции
Офтальмологические конференции и симпозиумы
Видео
Видео докладов

Поздравляем–> –> Онлайн трансляцииXIX Конгресс Российского глаукомного общества 3-4 Декабря 2021 9:00
В кору больших полушарий поступают афферентные импульсы от всех рецепторов организма. Непосредственной передаточной станцией этих импульсов к коре (за исключением импульсов, идущих от обонятельных рецепторов) являются ядра таламуса и прилежащих к нему образовании, где расположены третьи нейроны афферентных путей (стр. 542). Участки коры, в которые преимущественно поступают афферентные импульсы, И. П. Павлов назвал центральными отделами анализаторов.
| В | Центральные отделы, иначе корковое представительство, многих анализаторов, например кожного, суставно-мышечного (кинестетического), висцерального, пространственно совпадают и частично перекрывают друг друга. Области коры, в которых расположены центральные отделы анализаторов, принято называть сенсорными зонами коры больших полушарий (рис. 246).
Сенсорные зоны коры больших полушарий представляют собой корковую проекцию периферических рецепторных полей. Рис. 246. Локализация некоторых сенсорных и моторных зон в коре больших полушарий у человека (схематизировано). |
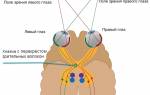
Представительство соматической и висцеральной чувствительности. В каждом полушарии имеются две зоны представительства соматической (кожной и суставно-мышечной) и висцеральной чувствительности, которые условно называются I и II соматосенсорными зонами коры. Первая соматосенсорная зона корыВ расположена в задней центральной извилине.
| В |
Размер ее значительно больше, чем второй. К этой зоне поступают афферентные импульсы от заднего вентрального ядра таламуса, доставляющие информацию, получаемую кожными (тактильными и температурным суставно-мышечными и висцеральными рецепторами противоположной стороныВ тела. На рис. 247 показано расположение в этой зоне проекций различных частей тела человека. Как видно, наибольшую площадь занимает корковое представительство рецепторов кисти рук, голосового аппарата и лица, наименьшую площадь — представительство туловища, бодра и голени. Рис. 247. Расположение в соматосенсорной зоне коры больших полушарий человека проекций различных частей тела (по У. Пенфилду н Расмуссену). 1 – половые органы; 2 – пальцы; 3 – ступня; 4 – голень; 7 – шея; 8 – голова; 9 – плечо; 10 – локтевой сустав; 11 – локоть; 12 – предплечье; 13 – 15 – мизинец; 17 – средний палец; 18 – указательный палец; 19 – большой палец; 21 – нос; 22 – лицо; 24 – зубы; 25 – нижняя губа; 26 – зубы, десны и челюсть; 27 – язык; 28 – глотка; 29 – внутренние органы. Размеры частей тела соответствуют размерам сенсорного представительства В |
Площадь корковой проекции определяется количеством нервных клеток коры, участвующих в восприятии раздражений от того или иного рецепторного поля. Чем количество клеток больше, тем более дифференцирован анализ периферических раздражений. Корковые проекции рецепторов висцеральных афферентных систем (пищеварительного тракта, выделительного аппарата, сердечно-сосудистой системы) расположены в области представительства кожных рецепторов соответствующих участков тела.
Вторая соматосенсорная зона расположена под роландовой бороздой и распространяется на верхний край сильвиевой борозды; афферентные импульсы в эту зону также поступают из заднего вентрального ядра таламуса.
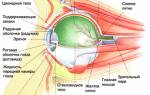
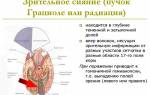
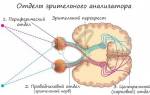
Представительство зрительной рецепции. Корковые концы зрительного анализатора, так называемые зрительные зоны, расположены на внутренней поверхности затылочных долей обоих полушарии в области шпорной борозды и прилегающих извилин. Зрительные зоны представляют собой проекцию сетчатки глаза. Афферентные импульсы поступают в эту область от наружных коленчатых тел, где находятся третьи нейроны зрительного пути.
Представительство слуховой рецепции. Корковые концы слухового анализатора локализуются в первой височной и так называемой поперечной височной извилинах Гешля. Афферентные импульсы поступают в эту зону от клеток внутренних коленчатых пути) и несут информацию от тел (третьи нейроны слухового тутовых рецепторов улитки внутреннего уха. Импульсы, возникающие в рецепторах улитки при восприятии тонов разной высоты, поступают в различные группы клеток слуховой зоны.
Представительство вкусовой рецепции. Корковые концы вкусового анализатора, по данным Пенфилда, расположены у человека в височной рядом с участком коры, ракздражение которого вызывает слюноотделение. Афферентные импульсы поступают во вкусовую зону от нижнего заднего ядра таламуса.
Представительство обонятельной рецепции. Пути обонятельной чувствительности являются единственными афферентными путями, не проходящими через ядра зрительных бугров. Их первые нейроны — обонятельные клетки — располагаются в слизистой оболочке носа. Вторые нейроны находятся в обонятельной луковице. Отростки вторых нейронов образуют обонятельный тракт, который доходит до клеток, расположенных в передней части грушевидной доли (Л. Бродал), где расположен корковый конец обонятельного анализатора.
Эффекты раздражения и разрушения сенсорных зон у человека. Локализация сенсорных зон у человека изучена главным образом методом электрического раздражения различных точек коры во время мозговых операций. Так как такие операции проводятся под местной анестезией, то пациент может дать точную словесную характеристику возникающих у него ощущений. Последние, как показали детальные исследования, проведенные Пенфилдом и др., всегда имеют элементарный характер. Так, при раздражении зрительной зоны у человека возникают ощущения вспышки света, темноты и различных цветов. Никаких сложных зрительных галлюцинаций при раздражении этой области не наблюдается. Раздражение слуховой области коры вызывает ощущения различных звуков, которые могут быть высокими и низкими, громкими и тихими; однако никогда при электрическом раздражении у пациента не возникает восприятия звуков речи. Раздражение соматосенсорной зоны вызывает ощущения прикосновения, покалывания, онемения, реже слабое температурное или болевое ощущение. Выраженных болевых ощущений почти никогда не наблюдается. При раздражении обонятельной или вкусовой зоны возникают различные запаховые или вкусовые (большей частью неприятные) ощущения.
Разрушение сенсорных зон у человека ведет обычно к грубым нарушениям данного вида чувствительности на противоположной очагу поражения стороне тела. Двустороннее поражение зрительных зон приводит к полной слепоте, удаление слуховых зон — к глухоте. Нарушения функций сенсорных зон у человека при кровоизлиянии, опухоли, ранении компенсируются значительно хуже, чем у животных. На основании опытов, проведенных на собаках с удалением разных участков коры больших полушарий. И. П. Павлов пришел к выводу, что в корковом конце каждого анализатора следует различать центральную часть, или ядро, и так называемые рассеянные элементы. Под этими элементами он понимал нервные клетки, расположенные в широкой области, куда поступают импульсы от тех же рецепторов, что и в ядре анализатора. Наличие рассеянных элементов обеспечивает возможность компенсации функции при разрушении ядра анализатора. У человека компенсация функций менее выражена, вероятно, потому, что нервные клетки корковых концов анализаторов более концентрированы в сенсорных зонах.
Регистр лекарственных средств России РЛС Пациент 2003. – Москва, Регистр Лекарственных Средств России, 2002.
Мы много говорили о регуляции функций клеток и организма. Теперь попытаемся сопоставить механизмы работы анализатора и регуляции функций организма, рассмотрев схему на рисунке 1.5.24. В ней обобщены механизмы работы органов и систем органов, по П.К. Анохину (теория функциональных систем) и К.В. Судакову (система регуляции артериального и осмотического давления). Проследив по рисунку за направлением светлых стрелок, можно отметить однонаправленность и последовательность процессов регуляции, а также замкнутость контура системы.
Рисунок 1.5.24. Схема информационной системы регуляции функций организма
Прежде чем рассматривать механизмы обработки информации, вспомним несколько определений и будем периодически возвращаться к этому рисунку. Анализатор (выделен на схеме темным фоном) – совокупность центральных и периферических образований, воспринимающих и анализирующих изменения окружающей среды и внутренней среды организма. По функциональному значению выделяют периферический (воспринимающий), проводниковый (передающий информацию) и центральный, или корковый (расположенный в коре полушарий головного мозга) отделы анализатора. Периферический отдел анализатора представлен рецепторами (рисунок 1.5.24 и 1.5.25). Отметим, что органы чувств (раздел 1.5.2.10) по сути – те же рецепторы, реагирующие на определенные внешние стимулы. Рецепторы улавливают изменения показателей текущего состояния, а затем проводят первичный анализ информации: сравнивают полученные значения с генетически запрограммированными, заданными как эталон (норма). Другими словами, они выдают информацию о разности текущего и заданного значений параметра. Специфичность рецепторов заключается в их способности воспринимать раздражитель только определенного вида. Например, зрительный рецептор воспринимает только электромагнитные волны с длиной волны 390-760 нанометров (свет, цвет), слуховые рецепторы – только звуковые волны в диапазоне от 16 Герц до 20 000 Герц, вкусовые и обонятельные рецепторы фиксируют химический состав пищи или вдыхаемого воздуха и так далее.
Как объяснить такую “узкую специализацию” органов чувств и рецепторов вообще? Согласно одной из теорий, вынужденные колебания светового луча порождают резонансные колебания только рецепторов сетчатки. Другие рецепторы (обоняния, вкуса, тактильные и другие) имеют иные частоты собственных колебаний и поэтому на световой луч не реагируют (не вступают в резонанс). В зависимости от источника стимулов (возмущающих воздействий), на которые реагирует рецептор, различают три группы рецепторов (рисунок 1.5.25).
Рисунок 1.5.25. Классификация рецепторов
Экстерорецепторы (от латинского exterus – находящийся снаружи, внешний) воспринимают раздражение извне: звук, свет, запах, вкус, прикосновение, боль, температуру окружающих предметов и другие характеристики. Проприорецепторы (от латинского proprius – собственный) улавливают изменения в состоянии опорно-двигательного аппарата. Они воспринимают сокращение мышечных волокон, натяжение сухожилий и связок, положение отдельных частей тела (согнута в колене нога, поднята вверх рука и так далее). Интерорецепторы (от латинского interior – внутренний) располагаются во внутренних органах, тканях, в стенках кровеносных и лимфатических сосудов. Интерорецепторы регистрируют изменение химического состава (например, уровень глюкозы в крови или соляной кислоты в желудочном соке), давление, механические и другие изменения характеристик внутренней среды организма.
Полученная информация через проводниковое звено анализатора посредством нервной, гормональной, метаболической и миогенной систем регуляции (рисунок 1.5.24) передается в центры головного мозга и, в частности, лобные доли коры полушарий, отвечающие за сознание. Информация анализируется, после чего вырабатывается управляющий сигнал, поступающий посредством нервных, гормональных, метаболических и миогенных механизмов к эффекторам. Эффекторы здорового организма под влиянием управляющего воздействия корригируют параметры текущего состояния, подстраивая их до нормальных (эталонных) значений, что фиксируется рецепторами. Круг замкнулся. Отметим, что внешние факторы могут оказывать воздействие (показано темными стрелками) на любое из звеньев механизма регуляции и влиять (негативно или позитивно) на результат – значение исследуемого параметра.
Как осуществляется процесс передачи информации? Как некий объект окружающей среды оставляет “отпечаток” в нашем сознании? Где хранится архив таких “отпечатков”? Попробуем ответить на эти вопросы.
В ответ на раздражение в рецепторах кодируется качественная характеристика раздражителя (свет, звук), сила, время и локализация его действия, а также месторасположение источника воздействия в окружающем пространстве. Например, свойства предмета (форма, цвет, размер, удаленность объекта), попавшего в наше поле зрения, в рецепторах сетчатки кодируются в результате фотохимических реакций, а затем перекодируются в электрический импульс нервного волокна. В проводниковом отделе анализатора при передаче сигнала от одного нейрона к другому происходит смена информационного кода. Поясним это на примере. Электрический сигнал мембранного потенциала при “переключении” в синапсе преобразуется в химический (выделение определенной “порции” медиатора) и наоборот, во втором нейроне химический сигнал вызывает электрический импульс. Таким образом, информация о внешнем стимуле многократно кодируется и перекодируется, пока сигнал возбуждения не достигнет центрального (коркового) отдела анализатора. Отметим, что смысловое значение передаваемого сигнала обычно остается прежним, и лишь при определенных негативных обстоятельствах содержание информации может искажаться в большей или меньшей степени.
|
Информация об объекте в процессе восприятия, передачи и обработки сигнала многократно кодируется и перекодируется. При этом смысловое значение информации обычно сохраняется. |
|
Для каждого раздражителя эволюция нашла свой оптимальный способ передачи информации. Для многих периферических нервных волокон была установлена логарифмическая зависимость между интенсивностью раздражителя и частотой вызываемых им вынужденных колебаний. Существует множество моделей, описывающих механизм передачи информации в нейронных сетях.
В процессе передачи информации могут принимать участие одновременно множество рецепторов и нервных волокон (рисунок 1.5.26). На роль ансамбля нейронов в кодировании и передаче информации в человеческом организме впервые указал Д. Хебб. Ансамбль нейронов – это группа нейронов, имеющая общий для них раздражитель. Д. Хебб высказал предположение, что информация передается исключительно через возбуждение группы нейронов, он предложил рассматривать ансамбль нейронов в качестве основного способа кодирования и передачи информации. Разные наборы возбужденных нейронов одного и того же ансамбля соответствуют разным параметрам раздражителя. То же происходит и с эфферентными (центробежными) сигналами. Разные наборы сигналов эфферентных звеньев соответствуют разным реакциям эффекторов. Такой способ передачи информации вполне надежен, так как не зависит от состояния одного нейрона.
Особый принцип обработки информации вытекает из детекторной теории. Он получил название принципа кодирования информации номером детектора (детекторного канала). Передача информации по номеру канала (термин предложен Е.Н. Соколовым) означает, что сигнал следует по цепочке нейронов, конечное звено которой представлено нейроном-детектором простых и сложных признаков, избирательно реагирующим на определенный физический признак или их комплекс. Такой подход присутствовал уже в опытах И.П. Павлова с кожными анализаторами собаки. Раздражение некоторого участка кожи вызывало очаг возбуждения в строго определенном участке коры головного мозга, отвечающий за соматические раздражения (раздражение участков тела).
Нейрон-детектор представляет вершину иерархически организованной нейронной сети (ансамбля нейронов). На более высоком уровне несколько нейронов сходятся на одном нейроне-детекторе, образуя его локальную нейронную сеть (рисунок 1.5.26). Величина возбуждения, возникающего в ансамбле нейронов и переданного на детектор, характеризуется силой возбуждения и его направленностью в виде суперпозициивозбуждений внутри ансамбля нейронов.
Несмотря на большие успехи, достигнутые при изучении нейронных процессов и функций различных групп нейронов, современная наука располагает значительно меньшими данными об интегративных механизмах мозга. Следует остановиться на понятии гностической единицы – нейроне более высокого порядка, на котором сходятся нейроны-детекторы. Возникновение в сознании целостного субъективного образа предмета связано с пирамидой последовательных нейронных преобразований, вершиной которой является операция, выполняемая гностической единицей (Е.Н. Соколов, 1996). Отдельным зрительным образам соответствуют отдельные гностические единицы. Они избирательно реагируют на сложные изображения (например, лица, жесты).
|
Иерархически организованная нейронная сеть организма сходится на гностических единицах. |
|
В итоге, поступающая в виде нервных импульсов информация перекодируется в структурные и биохимические реакции в гностических единицах. В коре головного мозга осуществляется высший анализ и синтез поступившей информации. Анализ заключается в том, что с помощью возникающих ощущений мы различаем действующие раздражители качественно (свет, звук, тепло и другие характеристики) и количественно (сила, продолжительность и локализация действия раздражителя), а также определяем месторасположение источника звука, света или запаха. Синтез реализуется в узнавании известного предмета, явления или в формировании образа впервые встречаемого предмета, явления. Узнавание явления или предмета в целом по совокупности восприятия отдельных характеристик раздражителя достигается в результате сличения поступающей в данный момент информации со следами памяти.
|
Без сличения ощущений со следами памяти узнавание невозможно. |
|
Если информация о предмете или явлении поступает в корковый отдел анализатора впервые, то формируется образ нового предмета, явления благодаря взаимодействию нескольких анализаторов. Но и при этом идет непрерывное сопоставление поступающей информации со следами памяти о других подобных предметах и явлениях. Поступившая в виде нервных импульсов информация кодируется в мозге с помощью механизмов долговременной памяти. Выделенные элементарные признаки синтезируются в долговременной памяти в передневентральной височной коре на гностических нейронах. Выполняется принцип:
|
Один образ – один гностический нейрон. |
|
Отображение воспринимаемого объекта в гностической единице характеризуется высокой степенью абстракции. При изменении размера, ориентации, цвета объекта гностический нейрон продолжает сохранять избирательность своей реакции на прежний объект. Осознание объекта в процессе его восприятия связывают с дополнительным свойством пирамиды образов. Оно реализуется механизмом синхронизации электрической активности мозга в диапазоне гамма-частот.
Итак, процесс передачи сенсорного сообщения сопровождается многократным перекодированием информации и завершается высшим анализом и синтезом, которые происходят в корковом отделе анализатора. После этого реализуется выбор или формирование алгоритмической программы ответной реакции организма.
Поделиться:
Опубликовано ср, 07/02/2018 – 15:46

Известно, что затылочная доля связана со зрением. Эта треугольная часть мозга расположена позади теменной доли. Медиальная граница затылочной доли ограничена теменно -затылочной бороздой. Латеральная сторона очерчена воображаемой линией, известной, как теменно – височная латеральная линия ( PO sulcus to the pre-occipital notch, parieto-temporal lateral line). Нижняя тенториальная поверхность очерчивается линией, соединяющей затылочную борозду с началом борозды PO ( теменно – затылочной ). Три поля Бродмана ( поля: 17, 18 и 19) также расположены в затылочной доле. На ее медиальной поверхности есть cuneus, извилина, расположенная между бороздой calcarine и PO sulcus. Лингвальная извилина отмечается на тенториальной поверхности затылочной доли (ниже по сравнению с бороздой calcarine); эта лингвальная извилина отделена от фузиформной извилины коллатеральной бороздкой. Фузиформная извилина очерчена латерально затылочно-височной бороздой.
Обнаружено положительная корреляция между лобной, височной, затылочной областями и эффективным решением тестов на коэффициент вербального интеллекта (IQ), устное обучение и различные исполнительные функции. Диагностическая синергия наблюдалась для альянса между вербальным IQ и правым височно-затылочным соединением и левой средней затылочной извилиной. В другом исследовании показано при этом увеличение активации в левой средней и верхней затылочной доле.
Затылочная эпилепсия не так распространена, как другие виды эпилепсии. Она широко известна среди клиницистов главным образом из-за ее редкости распространения (около 1,2-2,6% случаев среди всех случаев эпилепсии). Отличительными признаками затылочных припадков являются зрительные галлюцинации, однако они не присутствуют в каждом случае затылочной эпилепсии. Галлюцинации обычно инициируются в поле зрения контралатеральном к пораженной зрительной коре, а затем распространяются для охвата всего поля зрения. Преходящие визуальные симптомы, которые могут быть часто наблюдаемыми продуктивными симптомами (например, фосфены и вспышки), такжк, как и негативные симптомы, такие как амавроз, гемианопия и скотома, обычно описываются здесь пациентами. Статичные или мобильные вспышки, которые являются продуктивными симптомами, можно рассматривать как отличительную особенность вовлечения в патологический процесс затылочной доли. Визуальные галлюцинации становятся яркими и сложными, если имеет место вовлеченность патологического процесса в затылочно-височную кору, при этом наблюдаются галлюцинации со сценами различной сложности.
По сравнению с инфарктами других областей мозга, инфаркты затылочной доли различаются по этиологии, факторам риска и прогнозу. Пациенты с инфарктами затылочной доли, как правило, не имеют других неврологических симптомов, кроме потери зрения в определенном поле, и отличаются относительно легким течением, чем инфаркты у пациенов с многочисленными инфарктами или инфарктами в других областях головного мозга.
Пациенты с глубоким двусторонним затылочным повреждением жалуются на полную потерю сна. Более того, «минимальное распространенное поражение» ( ‘minimal lesion extension,’) , которое является обязательным ( облигатным ) для синдрома Шарко-Вилбранда ( Charcot-Wilbrand Syndrome – CWS), может быть представлено здесь при глубоком двустороннем повреждением затылочной доли в правой нижней лингвальной извилине. CWS может представлять собой дискретный и изолированный нейропсихологический симптом глубокого повреждения затылочных долей и может возникать даже в отсутствие фиксируемых аберраций сна REM и других профявлений когнитивного дефицита.
В контексте социального восприятия визуальная социальная информация проходит через вентральный поток во время первоначальной визуальной обработки, а затем направляется в такие области, как префронтальная кора, когда необходима когнитивная обработка более высокого порядка. Визуальная обработка лица происходит в вентральном тракте ( потоке), а положение активации зависит от задачи, указывая на иерархическое расположение вентрального потока. Таламус может включаться в визуальную обработку лица, вместе с височно-затылочной корой, фузиформной, пара цингулярной извилинами и лингвальной извилиной, в то время как стимулы лица и тела также активируют верхнюю височную борозду и среднюю затылочную извилину. Вентральный поток также может быть связан с восприятием эмоциональных раздражителей, благодаря значительной связи с миндалинами. Таким образом, первоначальная визуальная обработка, эмоциональная и социальная информация обрабатывается в вентральном потоке в сочетании с таламусом, височными и затылочными долями.
У пациентов с шизофренией наблюдалось уменьшение фракционной анизотропии внешнего белого белого вещества, рассеянного поперечно в каждом полушарии, особенно в левых височных и двусторонне в затылочных долях по сравнению с контрольными субъектами ( фракционная анизотропия различаются в зависимости от родства с пациентом как в полушариях, так и в затылочной и височной областях).
Существует достаточно доказательств, подтверждающих представление об уменьшении общего объема затылочной доли у пациентов с шизофренией. Краниальная асимметрия способна предсказать увеличение префронтальной маркировки у пациентов с шизофренией. Сравнение параметров микроструктурной диффузии относительно всей мозговой воксельной карте между шизофренией в ее дефицитарном (SZD) и не-дефицитарном подтипах (SZND) указывало на то, что радиальная диффузность была выше у левого форцепса у пациентов с SZD, в левой внутренней капсуле пациентов с SZND и в правом нижнем фронто – окципитальном фасцикуле во всей группе шизофрении. Средняя диффузность была увеличена от здорового контроля до пациентов с SZD и SZND в правой затылочной доле.
Длительная существование нелеченного психоза было связано с уменьшением объема серого височного и затылочно-височного серого вещества у больных с впервые лечившейся шизофренией. Также установлено, что у пациентов с шизофренией наблюдается значительно более низкая фракционная анизотропия в височном (верхнем височном и пара фунгиформном) и затылочном (верхнем и среднем затылочном) белом веществе по сравнению с пациентами с биполярным расстройством и здоровыми добровольцами. У некоторых больных шизофренией, отмечается увеличение плотности серого вещества в левой доле затылочной доле и базальных ганглиях. Некоторые авторы исследовали роль DTI белого вещества на ранней стадии шизофрении. Они обнаружили заметное снижение региональных значений функциональной анизотропии белого вещества в правой передней и правой средней частях затылочной доле.
У больных, страдающих шизофренией, уменьшается активность в средней затылочной извилине во время принятия решений. Существуют умеренные доказательства того , что снижение функциональной активности в правой лингвальной извилине во время кодирования эпизодической памяти и снижение активации фузиформной извилины отмечается при восстановлении эпизодической памяти у пациентов, страдающих шизофренией, по сравнению с контрольными группами.
Пациенты с шизофренией при визуализации их мозга демонстрируют снижение активации во время решения задач на обработку эмоций в лентиформной, фузиформной, средней окципитальной извилинах. Во время решения однозначных эмоциональных задач у пациентов с шизофренией наблюдается снижение активации в фузиформной извилине, а скрытая эмоция связана с уменьшением объема активности средней затылочной извилине. Также отмечаласьпри последующей активация памяти, активация в затылочном полюсе, латеральной затылочной коре, левой нижней височной извилине и фузиформной коре.
Патология теменно – затылочной и теменной области также выявлялись при шизофрении. При обработке задач на значения-аверсивных изображений по отношению к нейтральным изображениям, семейная группа риска продемонстрировали меньшую активацию передней части поясной извилины, с уменьшенным активности нижних префронтальной и затылочной части головного мозга. Снижение связей между цепочками нейронов (сцепления) было наиболее заметно между лобно-париетальными корковыми областями и подкорковыми структурами, а также между лобной и затылочной областями. Более того, усиленная связь была обнаружена между лимбической и фронтальной областями, а также для височно-затылочной связи.
Исследование, проведенное с помощью картографической статистики показало, что выраженность бреда связана с плотностью в орбитофронтальной коре (поле Бродмана 11/47). С другой стороны, бред, плоский аффект и несоответствие асоциального поведения с ситуацией были связаны с уменьшением плотности коры в дорсолатеральной префронтальной области. Hoptman et al., ( 2012) изучали функциональную связность между гомотопическими точками в головном мозге, используя гомосептическую связность воксель-зеркало (VMHC). Они провели исследование VMHC по данным fMRI В состоянии покоя и обнаружили заметные сокращения VMHC у пациентов в затылочной доле, таламусе и мозжечке.
У больных с ранним началом шизофрении было обнаружено атипичное снижение связи дорсолатеральной префронтальной коры с переднем цингулярном кортексом , нижней теменной долькой и средней затылочной извилиной. Fujimoto T et.al. ( 2012) провели исследование пациентов с шизофренией, собрав некоторые изображения в функциональной карте связанной с событиями дезинхронизации (ERD) и синхронизации (ERS) в диапазонах тета, нижнем альфа (8-10 Гц), верхнем альфа (10-13 Гц) и бета. Изменения ERD / ERS были отмечены в левом височном поле и средней линии фронтальной и передней коры головного мозга в тета-ERS, затылочной доле в альфа-ERS и правой височно-лобной – париетальной, средней линии фронтальной и передней коры головного мозга в бета-ERD.
Изучались волны дельты во время сна в двусторонних лобных, центральных, теменных и затылочных областях мозга. Было отмечено, что у пациентов с шизофренией наблюдалось более низкие показатели общего количества дельта-волн во время ночного сна.
Анализ психофизиологического взаимодействия дорзальной передней цингулярной коре показал, что активность в затылочной коре связана с плохим социальным изменением и нарушением социального познания ( Taylor S., et.al., 2011). Кроме того, fMRI показала три сети мозга, в которых имели место разрывы связей между нейронами у детей и подростков с шизофренией. Эти сети ( networks) включали переднюю цингулярную и височные доли, мозжечок с подкорковыми областями и затылочную долю и мозжечок. Пациенты с ранним дебютом шизофрении проявляли аномальную функциональную связность в сетях, касающихся трактов ( потоков )височной доли, лимбической области , мозжечка при ранней визуальной обработке. Кроме того, было обнаружено, что измененная префронто-гиппокампальная и парието-затылочная связь связана с позитивными симптомами шизофрении, что может иметь значение при анализе патофизиологии этих симптомов.
Пациенты, страдающие шизофренией, демонстрируют удивительно высокую плотность функционального связывания состояний (rsFCD) в двустороннем стриатуме и гиппокампе и значительно меньшее rsFCD в правой затылочной коре и двусторонней сенсомоторной коре. Уменьшается связь между левой нижней лобной извилиной с двусторонней затылочной корой у пациентов с шизофренией.
Уровни N-ацетиласпартата в соотношение с креатинином (NAA / Cr) , в левых базальных ганглиях и теменно-затылочной доле , было значительно ниже у пациентов с первым эпизод психоза шизофрении, чем у контрольных субъектов. Про – оксидантная ксантин – оксидаза (XO), которая, как установлено, играет неотъемлемую роль в метаболизме пурина / аденозина, снижает активность свою (XO) в затылочной коре и таламусе пациентов с психозом.
Итак, у пациентов с шизофренией наблюдаются структурные изменения (CT, МRI) в затылочной доле, а некоторые исследования показывают и функциональные изменения (fMRI, PET) вэтом регионе. С другой стороны, у некоторых пациентов наблюдаются метаболические изменения в затылочной доле. Умеренное доказательство качества (из-за большого размера выборки и отсутствия согласованности и точности) показало, что целостность белого вещества снижается в затылочной коре по сравнению со здоровыми индивидуумами, показания от умеренного до низкого качества (непоследовательные, неточные) показывают, что существует более высокая частота аномальная (обратная) асимметрия в затылочной доле при шизофрении по сравнению с контролем. Прогрессивные изменения объема серого вещества сообщались при продольном сканировании МРТ. Значительно большие сокращения сообщалось со временем при шизофрении по сравнению с контролем (затылочное серое вещество: 69,9%, а затылочное белое вещество – 45,9%).
Во время решения задач на исполнительное функционирование, больные шизофренией демонстрируют снижение активности в средней затылочной извилине.
Не вызывает сомнения , что затылочная доля связана с шизофренией, а аномалии затылочной доли могут быть важны для шизофрении и связанных с ней расстройств. Структурные нарушения мозга в теменной доле были обнаружены при исследованиях изображений при дебютах шизофрении в детском возрасте, предполагают, что аномалии серого вещества начинаются в теменных и затылочных долях и продолжают свою патологическую динамическую “волну” к фронтальной коре.
В мозге пациентов с шизофренией нет значительных метаболических изменений. Характер и сроки патологических процессов, лежащих в основе структурных, функциональных и метаболических изменений шизофрении, остаются неясными. Таким образом, структурные изменения в затылочной доле могут возникать на ранней стадии болезни и указывать на генетические влияния, которые еще предстоит определить. Нарушения в нейронных связей в цепях нейронов и трактов волокон , направляющихся к затылочной доле связаны с шизофренией и могут указывать на генетическую предрасположенность к этому психическому расстройству. В исследовании после смерти больных шизофренией Selemon et al. ( 1995) показали аномально высокую плотность нейронов в префронтальной и затылочной коре. Раньше смерть нейронных клеток объяснялась причиной уменьшения объема коры серого вещества. Однако, Selemon L, Goldman-Rakic P. ( 1997) связывают увеличение плотности клеток, в контексте уменьшенного объема, с потерей нейтрофилов или взаимосвязей между нейронами, что может быть результатом “чрезмерной обрезки” их отростков ( эксцессивного прунинга).
Категория сообщения в блог: Биологическая психиатрия